
Acute myeloid leukemia (AML) exhibits pronounced heterogeneity and chemotherapy resistance. Aberrant programmed cell death (PCD) implicated in AML pathogenesis suggests PCD-related signatures could serve as biomarkers to predict clinical outcomes and drug response. We utilized 13 PCD pathways, including apoptosis, pyroptosis, ferroptosis, autophagy, necroptosis, cuproptosis, parthanatos, entotic cell death, netotic cell death, lysosome-dependent cell death, alkaliptosis, oxeiptosis, and disulfidptosis to develop predictive models based on 73 machine learning combinations from 10 algorithms. Bulk RNA-sequencing, single-cell RNA-sequencing transcriptomic data, and matched clinicopathological information were obtained from the TCGA-AML, Tyner, and GSE37642-GPL96 cohorts. These datasets were leveraged to construct and validate the models. Additionally, in vitro experiments were conducted to substantiate the bioinformatics findings. The machine learning approach established a 6-gene pan-programmed cell death-related genes index (PPCDI) signature. Validation in two external cohorts showed high PPCDI associated with worse prognosis in AML patients. Incorporating PPCDI with clinical variables, we constructed several robust prognostic nomograms that accurately predicted prognosis of AML patients. Multi-omics analysis integrating bulk and single-cell transcriptomics revealed correlations between PPCDI and immunological features, delineating the immune microenvironment landscape in AML. Patients with high PPCDI exhibited resistance to conventional chemotherapy like doxorubicin but retained sensitivity to dasatinib and methotrexate (FDA-approved drugs for other leukemias), suggesting the potential of PPCDI to guide personalized therapy selection in AML. In summary, we developed a novel PPCDI model through comprehensive analysis of diverse programmed cell death pathways. This PPCDI signature demonstrates great potential in predicting clinical prognosis and drug sensitivity phenotypes in AML patients.

Acute myeloid leukemia (AML) arises from the abnormal clonal expansion of myeloid hematopoietic progenitor cells and represents a highly heterogeneous group of hematological malignancies 1 . Despite its low incidence, AML remains the most common type of acute leukemia in adults and the second most prevalent leukemia overall 2 . While novel treatment options continue to emerge, clinical outcomes of AML patients over 5 years remain disappointing, with many patients experiencing relapse and developing resistance to therapy, leading to treatment failure and mortality 3 . Prognosis in AML depends on several factors including disease history 4 , age 5 , and cytogenetic and molecular abnormalities such as mutations in NPM1, CEBPA, FLT3, and KIT 6 . Although incremental advancements have been made in the diagnosis and treatment of AML, favorable responses are often limited to a minority of patients, and the emergence of drug resistance frequently results in therapeutic failure, especially for elderly AML patients, with the majority succumbing to the disease within one year of diagnosis 1,3,5,7 . Therefore, the development of novel prognostic biomarkers to enable personalized risk stratification and prediction of therapeutic response is critically needed.
Programmed cell death (PCD), which enables the proper formation of organs during embryonic development, plays a fundamental role not only in animal development but also in the maintenance of tissue homeostasis 8,9,10,11,12 . Dysfunction of PCD is associated with multiple pathological conditions and the development of various diseases, including immune disorders, neurodegeneration, tissue damage, and cancers, such as breast cancer, liver cancer, and AML 9,10,13,14,15 . Currently, PCD consists of 13 major cell death patterns, including apoptosis, pyroptosis, ferroptosis, autophagy, necroptosis, cuproptosis, parthanatos, entotic cell death, netotic cell death, lysosome-dependent cell death (LDCD), alkaliptosis, oxeiptosis, and disulfidptosis 10,16,17 . Apoptosis is the most well-studied PCD pathway. Defective apoptosis is a hallmark of AML, contributing to treatment resistance 18 . Restoring apoptosis sensitivity in leukemia cells represents a promising therapeutic strategy 19 . Autophagy is a tightly regulated homeostatic process that controls cell survival and cell death. It involves the encapsulation of cytoplasmic contents within autophagosomes, which then fuse with lysosomes for degradation and recycling of the sequestered material 20 . Defects in autophagy can promote leukemogenesis in AML by enabling the survival of pre-leukemic cells 8 . Additionally, autophagy has been implicated in resistance to cytarabine 11 and targeted therapies like FLT3 in AML 21 . Disulfidptosis is a recently discovered iron-dependent form of programmed cell death induced by disulfide stress resulting from the accumulation of incorrect disulfide bonds within the endoplasmic reticulum 16 . While the role of disulfidptosis in AML pathogenesis remains to be fully elucidated, it may be involved in the perturbation of redox homeostasis in AML cells. Ferroptosis is an iron- and ROS-dependent form of PCD characterized by lipid peroxidation and loss of membrane integrity 17 . Aberrant expression of genes regulating ferroptosis, including downregulation of GPX4 and altered expression of NRF2, has been observed in AML, which may disrupt redox homeostasis 12,22 .
Other forms of PCD such as necroptosis, pyroptosis, lysosome-dependent cell death, and entotic cell death may also contribute to AML pathogenesis 23 . However, the association between the multiple PCD pathways as a whole and AML biology requires further study. Elucidating the roles of PCD-related genes could provide insight into AML pathogenesis and reveal potential therapeutic targets. In this study, we identified six pan-programmed cell death-related genes (PPCDS) demonstrating differential expression in AML. We subsequently developed a PPCDS index (PPCDI) to investigate potential associations between PPCDS and AML prognosis. The genetic and mutational landscape of these PPCDS in AML was characterized. Prognostic models incorporating PPCDS and PPCDI data were then constructed to predict overall survival (OS) in both training and validation AML cohorts. Furthermore, we examined interactions between PPCDS/PPCDI levels and immune features in AML. Additionally, the immunotherapy and targeted therapy response related to PPCDS/PPCDI status in AML was evaluated and verified.
RNA sequencing (RNA-seq) expression data (log2 transformed) were obtained for 129 AML patients from The Cancer Genome Atlas (TCGA) database (https://portal.gdc.com). Additionally, the expression profiling data for three external AML cohorts were acquired from the Tyner’s cohort 24 , GSE37642-GPL96 25 , and Target (https://ocg.cancer.gov/programs/target) datasets, respectively. Corresponding clinicopathological characteristics for AML patients from these three cohorts were also collected. Furthermore, single-cell RNA sequencing (scRNA-seq) data from 20 AML patient samples were retrieved from the GSE185993 dataset 26 .
A total of 3,246 PCD-related genes were compiled from the KEGG database, GSEA gene sets, and previous publications by integrating 13 major forms of PCD. These encompassed genes involved in apoptosis (n = 580), pyroptosis (n = 52), ferroptosis (n = 91), autophagy (n = 370), necroptosis (n = 104), cuproptosis (n = 14), parthanatos (n = 9), entotic cell death (n = 15), netotic cell death (n = 8), lysosome-dependent cell death (n = 220), alkaliptosis (n = 7), oxeiptosis (n = 5), and disulfidptosis (n = 1771), as categorized based on prior reports 10,17 (Table S1).
To further identify the PPCDS associated with prognosis in AML, 10 machine learning algorithms were implemented, including Supervised Principal Components (SuperPC), CoxBoost, stepwise Cox regression, Least Absolute Shrinkage and Selection Operator (LASSO), Generalized Boosted Regression Modeling (GBM), Survival Support Vector Machine (Survival-SVM), Partial Least Squares regression for Cox models (plsRcox), Ridge regression, Elastic Net (Enet), and Random Survival Forest (RSF). A total of 73 algorithm combinations were evaluated. To develop a robust PPCDS signature with optimal stability, the 73 models were trained in a leave-one-out cross-validation (LOOCV) framework 27 . Model performance was assessed using the concordance index (c-index), from which the final optimal algorithm combination was selected.
Consensus clustering (CC) analysis for the TCGA and Tyner’s cohort was performed utilizing the ConsensusClusterPlus R package 28 . The algorithm was configured for agglomerative hierarchical clustering by partitioning around medoids, with a Pearson correlation distance metric. Resampling of 80% of the sample population was conducted for a total of 1000 iterations. Principal component analysis (PCA) was implemented using the R stats package to normalize the expression data. This involved z-score calculation followed by dimensionality reduction through principal component extraction using the prcomp function. This generated an optimized low-dimensional matrix for subsequent analyses.
Based on PPCDI level, patients in the TCGA-AML cohort were divided into PPCDI-high and low subgroups, the mutation features of patients in various groups were explored using the maftools package. Additionally, we examined the alteration landscape of each PPCDS in AML based on 165 samples from the TCGA-AML dataset using cBioPortal database (with full expression, mutation, and CAN data) 29 .
Gene set enrichment analysis was conducted on the DEGs using the clusterProfiler and ggplot2 R packages. Enrichment for Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways and Gene Ontology (GO) biological process terms was evaluated. Additionally, the Gene Set Variation Analysis (GSVA) was utilized to assess differences in HALLMARK gene set functions between the high and low PPCDI groups across the three cohorts, referencing the h.all.v2022.1.Hs.symbols.gm database 30 .
Prognostic nomogram models incorporating PPCDI and relevant clinical variables were constructed to predict the clinical outcomes of AML patients across the three cohorts 31 . Model clinical utility was evaluated through calibration analysis, decision curve analysis (DCA), and time-dependent receiver operating characteristic (ROC) curve analysis implemented in the rms, survival, and timeROC R packages, respectively 32,33 .
Single-cell RNA sequencing data from 20 AML samples were obtained from the GEO database (GSE185993) and analyzed using the Seurat and SingleR packages 26 . Quality control and preprocessing were performed, involving data normalization, variable feature selection, scaling, dimensionality reduction with PCA, batch effect correction via Harmony, clustering with shared nearest neighbor modularity optimization, UMAP dimensionality reduction, and differential expression analysis to identify cluster markers. This yielded a total of 42,475 high quality cells for downstream analyses. SingleR analysis coupled with known literature-based markers were utilized to assign cell type labels 26 . Marker gene functional enrichment for each cluster was evaluated with the ClusterGVis 34 , org.Hs.eg.db and enrichCluster R packages. Copy number variation (CNV) scores were calculated for each proliferating cells using inferCNV algorithm, with T cells set as the reference 35 . Pseudotime trajectory analysis was conducted using the CytoTRACE 36 and Monocle2 37 packages. Furthermore, the PPCDS scores were calculated for each cell using the AddModuleScore function.
The expression profiles of PPCDS along with abundances of infiltrating immune cell populations in the tumor microenvironment were analyzed for TCGA-AML and Tyner AML cohorts. Estimation of immune cell type scores was performed by applying a series of computational methods including TIMER, QUANTISEQ, EPIC, MCPCOUNTER, and ESTIMATE algorithms through the IOBR package 38 .
To investigate potential associations between the PPCDS/PPCDI signature and immune checkpoint molecule expression in AML, six immune checkpoint-related transcripts were selected for analysis, including CD274, CTLA4, LAG3, PDCD1, TIGIT, and SIGLEC15. The expression profiles of these genes were extracted from both the TCGA-AML and Tyner AML cohorts. The Tumor Immune Dysfunction and Exclusion (TIDE) algorithm 39 was utilized to predict differential immunotherapy response between high and low PPCDI subgroups. Additionally, the “oncoPredict” R package 40 was leveraged to evaluate differences in drug sensitivity.
This investigation was sanctioned by the Ethics Committee of the First Affiliated Hospital of Anhui Medical University. The study was conducted in accordance with the protocol, the principles of the “Declaration of Helsinki” in its current version and the requirements of the Chinese law. The cohort comprised twelve patients with acute myeloid leukemia (AML) newly diagnosed at the institution’s Hematology Department, alongside twelve healthy adults ascertained from the hospital’s Health Examination Center, all of whom provided written informed consent. Peripheral blood specimens were the biological material of choice, with AML diagnoses aligning with the guidelines delineated for the management of adult acute myeloid leukemia in China, acute promyelocytic leukemia excepted.
The transcriptional profiles of genes within the PPCDI cluster were scrutinized by leveraging RT-qPCR analysis to compare the expression between AML and normative healthy samples. The extraction of total RNA was facilitated by the Fastagen RNA extraction kit, subsequent to which cDNA synthesis was executed utilizing the M-MLV reverse transcriptase kit (Takara, Japan), adhering strictly to the protocols prescribed by the manufacturer. Quantitative assessments were conducted employing the ABI PRISM 7500 Sequence Detection System (Applied Biosystems) in conjunction with a dedicated RT-qPCR reagent kit (Promega). Quantitative data were procured via the standard curve method and the expression values were normalized against those of the β-actin (human) gene, serving as an internal reference.
Statistical analyses were conducted using R software (v4.2.1). Between-group comparisons of two groups were performed using Student’s t-test or Wilcoxon rank sum test as appropriate, while Kruskal-Wallis one-way analysis of variance was applied for comparisons involving more than two groups. Kaplan-Meier survival curves were generated and differences between curves were assessed. Associations between continuous variables were evaluated using Spearman’s rank correlation analysis. A p-value threshold of 0.05 was set for statistical significance in all analyses.
This study was reviewed and approved by the First Affiliated Hospital Ethics Committee of Anhui Medical University. Ethics approval number 2023342. Before participating in the study, all patients and healthy volunteers provided written informed consent.
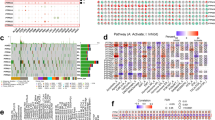
This study utilized 129 patients with complete clinical data from TCGA database as the training cohort. The validation cohorts consisted of 398 patients from the Tyner’s dataset and 417 patients from the GSE37642-GPL96 dataset. Clinical parameters for patients across the three cohorts are presented in Table S2, and the study workflow is outlined in the Fig. 1. To identify robust prognosis-associated DEGs, the intersection of DEGs from the three cohorts was derived, yielding 140 common DEGs (Fig. 2A,B, Tables S3–S5). Moreover, we performed a hypergeometric test to evaluate the overlap of the three datasets’ DEGs. The test yielded a p-value of 7.00e-49, demonstrating that the overlap is highly significant and unlikely to be due to statistical chance (Fig. 2B). GO and KEGG enrichment analysis revealed these 140 DEGs were highly enriched for pathways related to cell adhesion and lipid metabolism (Fig. 2C, Table S6). By overlaying the DEGs with genes associated with specific PCD forms, 30 pan-PCD related DEGs were identified, including 15 disulfidptosis, 1 pyroptosis, 2 ferroptosis, 3 lysosome-dependent cell death, 2 netotic cell death, 9 apoptosis, and 5 autophagy genes (Fig. S1, Table S7).


To identify optimal algorithms for constructing PPCDI, 73 machine learning models utilizing 10 different algorithms were evaluated. As shown in Fig. 2D, StepCox, RSF + StepCox, and LASSO + StepCox exhibited the highest average c-index (Table S8). To mitigate variable selection bias, the RSF and LASSO methods were intersected to derive a robust PPCDS signature, followed by multivariate Cox regression for final model building. The PPCDI score for each patient was calculated by summing the products of gene expression and corresponding coefficients. Patients were stratified into PPCDI-high and low subgroups based on the PPCDI median value. The RSF analysis identified 15 top-ranked genes by importance (Fig. 2E). By intersecting with the LASSO-selected genes, 6 core PPCDS genes were defined: ATP6V0E2, DDIT4, ETS2, DOCK1, CLEC11A, and HGF (Fig. 2F). Chromosomal locations and co-expression patterns were examined for each PPCDS (Fig. 2G). Most PPCDS demonstrated positive correlation in the TCGA-AML cohort, except for CLEC11A and HGF (Fig. 2G). Additionally, univariate Cox regression and Kaplan-Meier analyses were used to evaluate the associations between individual PPCDS and AML prognosis across cohorts. ATP6V0E2, DDIT4, ETS2, and DOCK1 conferred risk, while CLEC11A and HGF had protective effects (Fig. S2). Consistent results were observed in the Tyner’s and GSE37642-GPL96 cohorts (Fig. S3A,B). To ensure the accuracy of the above findings, we incorporated another outside cohort study involving 279 AML patients from Target datasets, analyzing the association between PPCDS and the clinical outcomes of AML patients. The findings indicated that the PPCDS displayed consistent behavior across all cohorts, further validating their potential as biomarkers (Fig. S3C).
Multivariate Cox regression analysis was conducted in the TCGA-AML cohort to determine the coefficients for each PPCDS gene in the PPCDI model: PPCDI = 0.27673 × ATP6V0E2exp + 0.099852 × DDIT4exp + 0.13156 × ETS2exp + 0.090291 × DOCK1exp-0.094115 × CLEC11Aexp-0.14942 × HGFexp. Of notice, elevated PPCDI significantly correlated with worse clinical outcomes and older patient age across all three AML cohorts (Fig. 3A–C). Using the PPCDS expression data, AML patients in the TCGA-AML and Tyner cohorts were clustered into two groups (C1 and C2) via consensus clustering (Fig. 3D,E). Kaplan-Meier analysis revealed patients in cluster C2 had improved OS compared to C1 (Fig. 3F,G, P < 0.001). Furthermore, alluvial diagram analysis demonstrated the majority of patients in cluster C1 had higher PPCDI scores and older age (Fig. 3H,I). The expression patterns and its association with the clinical features of AML patients in TCGA-AML cohort was shown in Fig. 3J.

Somatic mutation analysis was performed comparing AML patients stratified into high and low PPCDI groups. Common mutated genes identified included DNMT3A, RUNX1, and NPM1, with higher mutation frequencies observed in the PPCDI-high subgroup (Fig. S4A). To further investigate the genomic alteration landscape of individual PPCDS in AML, the cBioPortal database was analyzed. PPCDS exhibited low alteration frequencies overall, with ETS2 harboring the highest mutation rate at 10% (Fig. S4B).
Patients in the TCGA-AML, Tyner, and GSE37642-GPL96 cohorts were stratified into high and low PPCDI groups based on calculated PPCDI scores. Elevated PPCDI significantly associated with poor clinical outcomes of AML patients (Fig. 4A,B). Principal component analysis clearly separated the AML patients into PPCDI-high and low subgroups in each cohort (Fig. 4C). GSVA was subsequently performed to evaluate differences in HALLMARK pathway activity between PPCDI-defined subgroups in the Tyner cohort, revealing enriched pathways (Fig. 4D,E). Notably, cross-cohort analysis identified consistent enrichment of oncogenic pathways, including KRAS_SIGNALING_UP, IL2_STAT5_SIGNALING, P53_PATHWAY, and TGF_BETA_SIGNALING pathways (Fig. 4E, Table S9).


To examine PPCDS expression and distribution at single-cell resolution, scRNA-seq data from AML patient samples (GSE185993) were analyzed. After quality control, 42,475 cells were retained for downstream analysis (Fig. S6A). Clustering identified 28 subpopulations, which were annotated into 7 cell types (Fig. 6A, Fig. S6B). Marker gene expression profiles were generated for each cluster (Fig. 6B, Fig. S6C), with enrichment analysis of the top 5 biological processes. The expression and distribution patterns of PPCDS were shown in Fig. S6D. CNV scores were calculated for proliferative cells using the inferCNV algorithm, with T-cells as reference (Fig. 6C,D). This delineated CNV-low (clusters 11, 13) and CNV-high (clusters 1, 8, 15, 16, 19, 26) subsets (Fig. 6D). Pseudotime trajectory analysis via CytoTRACE and Monocle2 was then performed to predict differentiation trajectories of proliferative subpopulations. A clear lineage progression was observed from CNV-low to CNV-high states (Fig. 6E–G). PPCDS scores were calculated for each cell using AddModuleScore (Fig. S6E). Notably, PPCDS score significantly correlated with malignant status of proliferative cells, linking PPCDS expression to AML pathogenesis (Fig. 6H).

Additional analyses were conducted to identify differences in other relevant features between PPCDI-high and -low groups. First, associations were examined between PPCDS/PPCDI levels and the infiltrating of various immune cells calculated using various algorithms. Interestingly, ImmuneScore, ESTIMATEScore, and infiltration levels of CD4 + T cells, CD8 + T cells, NK cells, neutrophils, macrophages, and myeloid dendritic cells demonstrated significant positive correlations with PPCDI scores and most PPCDS genes including ATP6V0E2, DDIT4, ETS2, and DOCK1. Conversely, these immune features negatively correlated with CLEC11A and HGF expression in both TCGA-AML and Tyner cohorts (Fig. 7A, Tables S10, 11). Next, expression of six immune checkpoint molecules was analyzed, revealing predominantly positive associations between checkpoint genes and PPCDS/PPCDI levels (Fig. 7B,C, Tables S10, 11). Strikingly, PPCDS and PPCDI also positively correlated with TIDE and dysfunction scores in AML, implying potential superiority of immunotherapy responses in PPCDI-low patients (Fig. 7B).

To elucidate the relationship between the model and drug sensitivity, we calculated the half maximal inhibitory concentration (IC50) values for each drug in TCGA-AML samples, aiming to identify drugs with significant differential activity. The landscape of correlations and significance between drug sensitivities and PPCDI is presented in Fig. 7D and Table S12. We observed that the IC50 of doxorubicin was higher, while the IC50 values of dasatinib and methotrexate were lower in the PPCDI-high group (Fig. 7D–G). These findings imply that AML patients with elevated PPCDI levels tend to be resistant to standard chemotherapy regimens, yet may remain sensitive to two other FDA-approved drugs originally developed for different leukemias. Therefore, dasatinib and methotrexate represent promising options for salvage therapy in chemotherapy-resistant AML. The associations between model genes and established AML therapeutic targets are presented in Fig. 7E.
RT-qPCR analysis was employed to ascertain the disparities in the expression of PPCDS mRNA between cohorts of normal healthy individuals and those with newly diagnosed AML. The sequences of the PCR primers targeting PPCDS are delineated in Table S13. As illustrated in Fig. 8A–D, a marked upregulation in the expression levels of ATP6V0E2, ETS2, DOCK1, and DDIT4 mRNA was observed in the AML cellular samples relative to the normal counterparts, corroborating the insights gleaned from bioinformatics analyses. Although the expression levels of CLEC11A and HGF did not show significant changes in the tumor cohort (ns, P > 0.05), as depicted in Fig. 8E,F, this observation is inconsistent with predictions from bioinformatics analyses, indicating that further elucidation of their expression at the protein level is warranted.

To the best of our knowledge, this is the first study to comprehensively investigate 13 diverse programmed cell death pathways, identify six PPCDS in AML, and construct a PPCDI signature using the TCGA-AML cohort. Validation of the PPCDI signature in two external AML cohorts (Tyner’s cohort, GSE37642-GPL96) confirms its performance. To evaluate the clinical utility of PPCDS in AML, we developed a nomogram incorporating PPCDI and relevant clinical factors, and verified its predictive applicability. Importantly, our findings reveal significant associations between PPCDI and the tumor microenvironment and drug sensitivity in AML. These results underline the potential value of PPCDI for stratifying AML patients and guiding personalized therapy.
PCD comprises biological and pathological processes that regulate tissue homeostasis, stress response, and cell proliferation, which is intricately associated with anti-tumor immunity in various cancers 9,10,13,17,20 . While prior studies have described roles for discrete PCD pathways in some cancers 10,14 , the genetic landscape, multimolecular mechanisms, and immune functions of pan-PCD genes in AML remain poorly characterized. Herein, we constructed a PPCDI signature based on six key PPCDS-ATP6V0E2, DDIT4, ETS2, DOCK1, CLEC11A, and HGF. We demonstrate this PPCDI signature can predict prognosis and therapy response in individual AML patients.
In a range of cancer types, the genes ATP6V0E2, DDIT4, ETS2, DOCK1, CLEC11A, and HGF have been subjected to extensive research. However, their functional roles and the mechanisms by which they are regulated within the context of AML remain insufficiently characterized. To corroborate the insights gleaned from our bioinformatics analysis, RT-qPCR was employed to assess the expression levels of PPCDS in samples from both healthy individuals and those diagnosed with AML. The findings largely corroborated the anticipated patterns of expression. Nonetheless, the levels of HGF and CLEC11A did not align with the bioinformatic predictions, signaling the necessity for further validation at the protein level.
ATP6V0E2 encodes a component of vacuolar ATPase (V-ATPase) that acidifies intracellular compartments. As mentioned previously, anlotinib can upregulate ATP6V0E2 expression and enhance lysosomal activity in colon cancer cells, resulting in attenuated apoptosis 41 . Of interest, AML cells also depend on lysosomal function to sustain redox homeostasis and survival. As such, increased ATP6V0E2 expression and lysosomal activation in AML could similarly protect leukemia cells from drug-induced apoptosis. DDIT4 is a stress-responsive protein that participates in mTOR pathway regulation, which is induced by various cellular stresses including DNA damage, oxidative stress, and nutrient deprivation 42 . Multiple studies have found high expression of DDIT4 associated with poor prognosis in AML patients 43 . DDIT4 may promote AML cell survival under chemotherapy by modulating autophagy and mTOR activity. ETS2 encodes an ETS family transcription factor that regulates gene expression and participates in the RAS/MAPK cascade 44 . Stromal ETS2 in the mouse pancreas helps recruit immunosuppressive cells like myeloid-derived suppressor cells and T regulatory cells, as well as chemokines 45 . Intriguingly, ETS2 may play a role in thymocyte proliferation and survival through a Myc-dependent pathway 46 . DOCK1 is a guanine nucleotide exchange factor that modulates Rho family GTPase signaling, which is critical for multiple cellular process, such as cell adhesion and migration, and actin cytoskeleton 47 . Specifically, the Dock1/Rac pathway is specifically targetable in the Npm1 cA/+ ; Smc3 Δ/+ AML with concurrent NPM1cA and cohesin mutations. Targeting this pathway could benefit this subtype, demonstrating the potential of genetics-guided precision oncology 48 . Conversely, upregulation of CLEC11A and HGF correlates with improved AML prognosis through various mechanisms 49,50 , consistent with our findings.
In this study, we did not observe significant associations between mutations in FLT3, IDH1, RAS, and NPMc and overall survival in TCGA-AML patients. The relatively low frequency of alterations in these individual genes within the cohort may partially account for the lack of detectable survival correlations. Moreover, AML is recognized as a high-risk disease driven by co-occurring combinations of mutations across many genes. Individual mutations alone may not exert sufficient prognostic influence 48 . In this study, we found upregulation of ATP6V0E2, DDIT4, ETS2, DOCK1 and downregulation of CLEC11A and HGF associated with poorer AML survival. A PPCDI signature calculated via multivariate Cox regression demonstrated robust prognostic performance, suggesting the PPCDI could be a potential prognostic biomarker in AML patients.
Increasing evidence indicates that tumor-microenvironment interactions influence T cell infiltration and immune cell function, relevant to immunotherapy 27,39 . We observed strong positive PPCDS/PPCDI correlations with immune infiltrates, indicating genetic-immunological crosstalk. High PPCDI patients exhibited elevated immune checkpoint molecules, implying heightened evasion and immunosuppression. Consistently, high PPCDI associated with increased TIDE and dysfunction scores, suggesting immunotherapy resistance. Moreover, elevated PPCDI associated with increased doxorubicin IC50 values, which may account for the poor prognosis resulting from chemotherapy resistance. In contrast, the IC50 values of dasatinib and methotrexate was lower in the PPCDI-high group, suggesting these agents as potential therapeutic alternatives for refractory AML.
Despite the potential of our model, some limitations warrant acknowledgement. First, the retrospective patient recruitment may have introduced partial bias. Second, the clinical utility of this model lacks validation in phase 3 randomized controlled trials, which are essential to inform treatment decisions. Third, due to the lack of IC50 data for other commonly used AML chemotherapeutic agents such as daunorubicin, cytarabine, mitoxantrone, and etoposide in the GDSC database, we were unable to investigate the association between PPCDI and the sensitivity of AML cells to these conventional chemotherapeutic drugs. Consequently, further investigations are warranted to assess the responsiveness of PPCDI to these standard AML chemotherapeutic agents. Although our study provides some preliminary evidence through in vitro PCR experiments, these results present limitations. To further validate the role and function of PPCDS in AML, more extensive protein level and functional experiments, such as Western blotting, co-immunoprecipitation, and cellular functional assays are required to comprehensively elucidate the specific mechanisms and roles of PPCDS.
In summary, we developed a six-gene pan-programmed cell death signature that provides novel insights into the interrelationships between cell death pathways and the clinical, molecular, and immunological landscapes of AML. Additionally, we constructed a robust PPCDS-based model capable of effectively predicting prognosis and responses to immunotherapy and targeted therapy in AML patients. Our findings indicate the potential clinical utility of PPCDS and PPCDI for prognostication and guiding personalized therapeutic decisions in AML patients undergoing immunotherapy and targeted therapies.
All data generated or analysed during this study are included in this published article [and its supplementary information files].